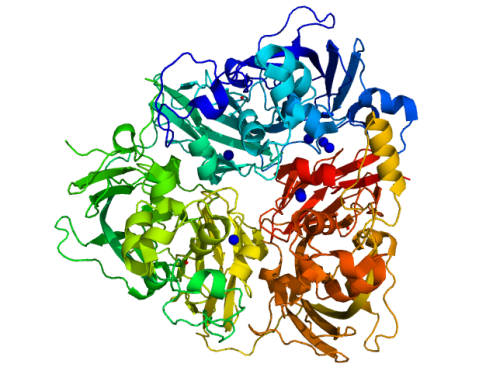
Вернемся к церулоплазмину человека. В крови человека и млекопитающих церулоплазмин фунционирует как неугомонный и неутомимый чистильщик, МЧС-ликвидатор и дезинфектор – молекулярный пылесос /molecular vacuum cleaner, выводя из обращения токсичные кислородные радикалы, окисляя ионы двухвалентного железа + ядовитые продукты “острой фазы” (при инфекциях) и гормоны стресса, адреналин, норадреналин и серотонин. См. также мой пост от 25 февр. 2020, где функция белка описана подробней. По своей молекулярной 3D-структуре это полноправный член эволюционного семейства 3-х и 6-ти доменных медьсодержащих оксидаз, которое включает церулоплазмин, гефестин, циклопен и примкнувшего к ним Шепилова – фактор коагуляции крови VIII (об этом мерзавце как-нибудь позже). Гефестин – это родной брат, почти структурный близнец церулоплазмина. В отличие от церулоплазмина, который “на все руки мастер” в русле крови, гефестин – белок внутриклеточный, целенаправленно работающий только на окисление токсичных ионов двухвалентного железа в клеточных факториях стенки кишечника.
Чтобы сотворить эдакое чудо, как церулоплазмин/ гефестин, природе нужно было приложить некоторые усилия и на каком-то этапе научиться соображать, думать то есть… Природе при родах церулоплазмина и гефестина нужно было понять, что самый лучший стройматериал для нашей большой структуры – это купредоксин (очень надежен, проверен в деле). Но купредоксин нам придется как-то ушестерить, т.е. купредоксин х 6 = церулоплазмин, см. картинку выше.
Ушестерение, это как? Попробуем ген купредоксина вначале дуплицировать, а затем этот дуплекс-прародитель триплицировать. Трудно, но в принципе, возможно. Дайте нам пару-тройку сотен миллионов лет, что-нибудь придумаем!
Да вот, еще вспомнили… Нам ведь нужно этим церулоплазмином что-то там окислять, пылесосить, ликвидировать и обезвреживать. А тут нам не обойтись без меди и аппарата Гольджи. Медь- Гольджи -медь, ау! * Давай-ка их сюда, в нашу коллекцию!
Все эти чудеса природы по сотворению белка церулоплазмина/ гефестина – намек ( в меру прозрачный) на заранее запланированную акцию, на присутствие невидимого дизайнера.
Честно признаемся: мысль о невидимом ‘телеологическом’ дизайнере – это нечто вынужденное, от бессилия найти рациональное объяснение.
А вообще-то здесь мы имеем дело с пока еще до конца не решенной загадкой структурной биологии.
Выживание наиболее эффективных и приспособленных молекул в процессе эволюции, дарвиновский селективный отбор – это, конечно, привлекательная модель. Однако, тут мы наталкиваемся на проблему времени, необходимого для случайного перебора вариантов конформаций молекул, а оно может быть бесконечно большим, соизмеримым со временем существования вселенной. См. например, мой пост от 5 марта 2020 о парадоксе Левинталя .
—–
*) Ядовитые субстраты и проч. молекулярная нечисть окисляются церулоплазмином в медьсодержащих активных центрах белка. Выбор ионов одно- и двухвалентной меди для вспомоществования в реакциях окисления в активном центре – это хороший, разумный выбор природы. Природа как бы почувствовала и просекла, что медь – это хорошо! Она увидела, что строение электронных оболочек меди, находящейся в группе переходных металлов 4-го периода таблицы Менделеева (там медленно заполняются d-оболочки от скандия -через железо и медь – до цинка) , это строение как нельзя более подходит для окисления широкого круга/ спектра органических и неорганических субстратов, веществ ядовитых и не очень. Встраивание ионов меди в молекулу церулоплазмина происходит на этапе посттрансляционной модификации в аппарате Гольджи сразу же после синтеза белка на рибосоме клетки.

Leave a comment